

Forebrain > Diencephalon> Epithalamus> Habenulae

Schematic showing the position of the habenulae in sagittal, horizontal and coronal sections through the zebrafish brain.
Based on the anatomical segmentation of 3 day old zebrafish larval brain by Thomas Müller, Olaf Ronneberger, Wolfgang Driever and colleagues. For details see Ronneberger et al., Nat. Meth. 2012 and http://vibez.informatik.uni-freiburg.de
Abbreviations: Ce, cerebellar plate; D, dorsal telencephalon/pallium; E, epiphysis; EmT, eminentia thalami; Hb, habenula; Hyp, hypothalamus; MO, medulla oblongata; OB, olfactory bulb; OT, optic tectum; PG, preglomerular complex; PO, preoptic area;PrT, pretectum; PTd, posterior tuberculum dorsal part; PTh, prethalamus; PTv posterior tuberculum ventral part; Teg, tegmentum; Th, thalamus; TS, torus semicircularis; V, ventral telencephalon/subpallium; Va, valvula cerebelli.
Introduction
The habenulae are part of a highly conserved conduction system, the dorsal-diencephalic conduction system (DDC), that interconnects sites in the limbic forebrain with the ventral midbrain and hindbrain (Bianco & Wilson, 2009).
The paired habenula nuclei exhibit several left-right asymmetries. Displaying differences in the proportion of neuronal subtypes with distinct patterns of gene expression, axon terminal morphology and connectivity (Bianco et al, 2008, Roussigne at al, 2009).
The habenulae have been shown to be important in a range of behaviours. Their regulation of monaminergic activity in the midbrain is important for the modulation of fear behaviours, avoidance learning and attention (Agetsuma et al, 2010). As would be expected of a regulator of monoaminergic activity they also have been implicated in psychosis, depression, anxiety, schizophrenia and addictive behaviours. They also play a role in circadian rhythms, sleep initiation and duration and reproductive and maternal behaviours (Bianco & Wilson, 2009, Hikosaka, 2010).
The neuroanatomical asymmetries of the habenula can also lead to a lateralisation of certain behaviours (Barth et al, 2005).
Neuroanatomy
The habenulae are spheroid nuclei. Habenular neurons are mono-polar.. The cells direct their dendrites inwards forming an internal neuropil core that the cell bodies surround.

Single-depth confocal section through the habenulae of a 4 dpf Tg(ET16:GFP) transgenic larva in which a subset of habenular neurons express GFP. In each nucleus the neuronal somata are arranged as ovoid shells surrounding a central neuropil domain. It is in this domain that neurons elaborate their dendritic arbors(Image courtesy of Isaac Bianco taken from Bianco et al, 2008).

A Left habenular neuron visualised by focal electroporation with a construct driving the expression of membrane GFP. Habenular cells have a rounded nucleus and are mono polar, projecting their dendrites inwards to form the dense neuropil of the habenulae (Image courtesy of Isaac Bianco taken from Bianco et al, 2008).
In the zebrafish, the left habenula receives projections from the left-sided parapineal organ that promotes the elaboration of left sided habenular identity (Bianco et al, 2008).
At later stages, in both habenulae a dorsal and a ventral habenular subnucleus(vHb) can be distinguished. The ventral subnucleus is homologous to the lateral mammalian habenula and the dorsal habenula is homologous to the medial mammalian habenula. The dorsal habenula can be further subdivided into lateral and medial subnuclei (dHbL,dHBM). (Amo et al, 2010).

A dorsal view of the epithalamus showing the left sided parapineal nucleus innervating the left habenular. The left-sided parapineal organ that promotes the elaboration of left sided habenular identity (Image courtesy of Miguel Concha, Concha et al, 2003).
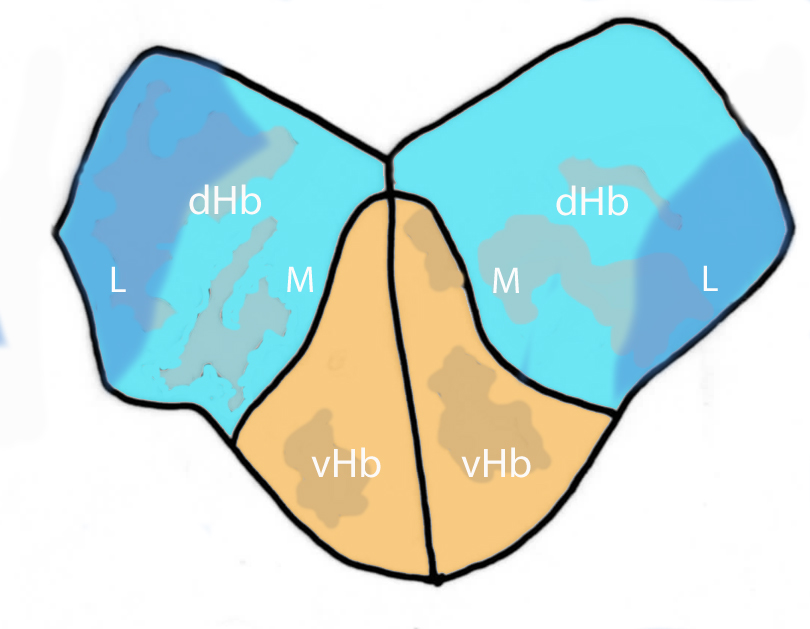
Schematic showing a transverse section through the adult habenular nuclei. This schematic shows the arrangment of the dorsal and ventral habenular subnuclei, the dorsal nucleus is further subdivided into medial and lateral subnuclei (See Development section to see the morphogenetic movements that result in this adult arrangement)(Schematic adapted from Amo et al, 2010).

This Figure shows a dorsal view of a 4dpf Tg(mp558b:GFP) embryo labelled with anti-acetylated tubulin(red) and SV2(blue). The anti-acetylated tubulin antibody labels axonal tracts and the SV2 antibody labels synaptic neuropil. This line expresses GFP only in the left dHbL nuclei. In panels B & C the same confocal stack has been rendered into 3D. By rotating the stack to the right we can see the synaptic neuropil of the three subnuclei and their relative positions at this stage of development. In panel C, the region that would be GFP positive is indicated by the dashed white line.
The habenulae are part of a system that relays information from the telencephalon and entopeduncular nucleus in the forebrain to the interpeduncular nucleus and to the raphe nucleus located in the midbrain/hindbrain. The afferent fibres from the telencephalon/ entopeduncular nucleus reach the habenulae via the stria medullaris. The projections of the habenula to the interpeduncular nucleus and raphe nucleus travel in the fasciculus retroflexus.
In zebrafish there are obvious anatomical asymmetries between the left and right habenulae. This can be seen in the differing size and shape of the neuropil in the left and right habenulae. The dHbM and dHbL subnuclei also have different size ratios on the left and the right and innervate different regions of the IPN. While the dHbm subnucleus is the main source of projections to the ventral region of the IPN the dHbL subnucleus projects mainly to the dorsal IPN (Agetsuma et al, 2010).

Dorsal view of a 4dpf wt embryo labelled with sytox nucleic acid stain, anti-acetylated tubulin(red) and SV2(blue). The spheroid nuclei of the habenulae can be seen surrounding a dense core of neuropil that is larger in the left habenular than in the right and differently shaped.
Left-right asymmetries in the telencephalon-habenula-IPN pathway.
(a) Schematic of a 4 dpf larval zebrafish brain. The habenular (Hb) nuclei receive afferent inputs from nuclei in the telencephalon (Tel) via the stria medullaris and diencephalon (not shown). Habenular neurons send efferent projections via the fasciculus retroflexus (FR) to the interpeduncular nucleus (IPN) in the ventral mid-brain. The L habenula predominantly innervates the dIPN and makes less substantial projections to the vIPN whereas the vast majority of R-sided axons terminate in the vIPN. In addition to the habenulae, the epithalamus contains the pineal complex, comprising the photoreceptive pineal (P) and parapineal (pp) nuclei. The parapineal is located on the L side of the dorsal midline and projects efferent axons that exclusively innervate the L habenula. (b) Neuroanatomical asymmetries in the epithalamus. Anti-acetylated tubulin immunostaining (red) reveals the L habenula displays a greater density of neuropil, especially in the dorso-medial aspect of the nucleus (arrow). The pineal (blue) and parapineal (green) nuclei are visualized by expression of GFP in a Tg(foxD3:GFP) transgenic larva. Parapineal efferent axons predominantly terminate in the asymmetric neuropil of the L habenula. (c) Three-dimensional reconstruction of habenular axon terminals in the ventral midbrain. L-sided axons were labelled with DiD (red) and R-sided axons with DiI (green). The dIPN is almost exclusively innervated by L-sided axons, whereas the ventral target receives a majority of R-sided inputs. All panels show dorsal views, anterior top. pc, posterior commissure;
Single cell electroporations of habenula neurons showed that habenular efferents have two distinct patterns of axonal arborisation in their terminals in the IPN. Axons coming from the left-sided habenula neurons terminate predominantly in the dorsal IPN. Three-dimensional reconstructions of these L-typical arbors reveal them to be formed like a 'domed crown' with branches extending over considerable DV depth. Their terminals circle the IPN and extend colaterals directed internally into the IPN.Right-sided habenula neurons predominantly terminate in the ventral IPN. Their axon terminals also surround the IPN but in a more ovoid-like shape “reminiscent of an electromagnetic coil. They look a lot flatter than the L-typical arbors, extending over less depth. The branches of the right axons concentrate more in the periphery, with relatively few branches in the centre of the IPN. Although these single cell electroporation experiments have not yet been carried out in transgenic lines labeling specific habenula subnuclei the results of Bianco et al, 2008 correlate well with later studies describing the innervation of the IPN by the lateral and medial dorsal habenula nuclei.

Habenular neurons have a stereotypical unipolar morphology and their axons terminate in spiral-shaped arbors that display multiple midline crossing. (Left) Three-dimensional reconstruction showing a R habenular (rHb) projection neuron in an intact 4 dpf larval zebrafish brain. Arrow indicates direction of axonal projection within the FR, from the rHb to the IPN. (Centre and Right) Confocal z-projection within the IPN showing a single axonal arbor elaborated by a L habenular projection neuron(centre) labeled by focal electroporation. The arbor crosses the ventral midline (dotted line) multiple times. Branches can also reverse direction such that they encircle the IPN in opposite senses. These arbors are shaped like a domed crown and arborize over a considerable DV extent. (Right) Shows an example of a R typical arbour, Which are considerably flatter. (Image provided by Isaac Bianco taken from Bianco et al, 2008)
From bulk labeling experiments, results suggest that the left habenular nucleus innervates the dorsal IPN and the right habenular nucleus innervates the ventral IPN. In Agetsuma et al, 2010 the habenula-IPN projection pattern in the adult is shown. Instead of bulk labeling the entire habenula nuclei with DiI, the dHbL and dHbm and their efferents are labeled in the double transgenic Tg(narp:GAL4VP16; UAS:DsRed2; brn3a- hsp70:GFP) fish. In the adult the Narp expression selectively labels the dHbL subnucleus, which is much larger on the left-side. Axons from this subnucleus terminate predominantly in the dorsal IPN. The Brn3a:GFP expression labels the dHbM, the terminals of these nuclei innervate the ventral IPN primarily. In the adult there is also an intermediate zone of the IPN where in the Tg(narp:GAL4VP16; UAS:DsRed2; brn3a- hsp70:GFP) red and green terminals overlap. The application of anterograde tracers to the dIPN showed that efferents from this region project to the dorsal raphé and continue on to terminate in the griseum central (GC)which corresponds to the mammalian periaqueductal gray (PAG), dorsal tegmental nucleus and nucleus incertus. These areas in mammals have been linked to the control of behaviour in fearful or stressful conditions. Manipulation of the dHbL-dIPN pathway with tetanus toxin light chain biased the fish towards a freezing behaviour as opposed to the normal flight response when subjected to cued fear-conditioning. This corroborates that in zebrafish the dHbL-dIPN –GC pathway, as in mammals, is involved in the modification of behavioral responses in an experience-dependent manner (Agetsuma et al, 2010).
Unlike the dorsal habenular nuclei the ventral habenular nuclei are symmetric, the vHb projects via the fasiculus retroflexus(FR) to the ventral part of the median raphé. The efferents travelling in the FR are arranged with the dHb efferents forming the core of the tract while the vHb efferents form a sheath surrounding them(Amo et al, 2010).
Habenula Afferent areas

Schematics showing a summary of the main afferent areas to the habenula from Turner et al., 2016.
(K: outline of the brain in lateral view, L: boxes diagram). (K) The brain nuclei that project to the habenula are shown as colored areas. (L) Boxes represent main nuclei projecting to the habenula, while arrows represent projections. Asymmetric projection from the parapineal (PP) to left habenula (lHB) is shown with a blue arrow. Asymmetric projection from mitral cells of the olfactory bulbs (OB) to right habenula (rHB) is shown with a pink arrow. All other afferent areas that seem to innervate both left and right habenula are represented with black arrows.
Development
By observing the expression patern of the vHB specific marker Diamine Oxidase(dao) throughout development it is possible to see that the dorso-ventral orientation of the fish habenula is a result of a morphogenetic process. At 5dpf expression of dao in the primordium is lateral to the dorsal habenular nuclei. This region then migrates ventro-medially to result in the arrangement we observe in the adult zebrafish.

Schematic showing the relative positions of the dosal and ventral habenulae subnuclei at different stages of development.
At 5dpf the vHb subnucleus is lateral to the dorsal habenular nuclei in the habenular primordium. This region then migrates ventro-medially to result in the arrangement we observe in the adult zebrafish. dHb (blue) and vHb(orange)(Schematic adapted from Amo et al, 2010).
Ontology
is part of: epithalamus, diencephalon, forebrain
has parts: dorsal habenula nucleus, ventral habenula nucleus
Transgenic Lines that label this brain region






Antibodies that label this brain region
Key Publications
Amo et al, 2010 http://www.ncbi.nlm.nih.gov/pubmed/20107084.
Concha et al, 2003 http://www.ncbi.nlm.nih.gov/pubmed/12895418
Bianco et al, 2008 http://www.ncbi.nlm.nih.gov/pubmed/18377638
Bianco & Wilson, 2009 http://www.ncbi.nlm.nih.gov/pubmed/19064356
Hendricks & Jesuthasan 2007 http://www.ncbi.nlm.nih.gov/pubmed/17394162
Agetsuma et al, 2010 http://www.ncbi.nlm.nih.gov/pubmed/20935642
Hikosaka, 2010 http://www.ncbi.nlm.nih.gov/pubmed/20559337
Turner et al, 2016 https://www.frontiersin.org/articles/10.3389/fncir.2016.00030/full
Back to Forebrain