
Anukampa Barth and Steve Wilson
Original paper reference
fsi Zebrafish Show Concordant Reversal of Laterality of Viscera, Neuroanatomy, and a Subset of Behavioral Responses
Although our body looks quite symmetrical on the outside, the localisation of many internal organs such as the heart and liver is asymmetric. Moreover, our brain shows anatomical and functional lateralization. Although this has been known for more than a century, until about 20 years ago our understanding of the differences between the left and right side of the brain was gathered mainly from post-mortem analyses and patients with specific brain lesions (such as strokes affecting one side of the brain). More recently, functional MRI studies in humans and experiments in animal models have helped to reveal more about the relationship between neuroanatomical structures and their function. Not only are our brains asymmetric in terms of structure and activity, some behaviours are also lateralised. An example is handedness in humans, and eye preference for detecting prey or predators in fish and amphibians.
In this publication, we show how we can use zebrafish to study brain asymmetries and how these asymmetries might relate to lateralised behaviours. We made use of a line of fish called frequent-situs-inversus (fsi), in which a proportion of the embryos show a reversal of the asymmetric placement of the viscera. For example, the direction of the looping of the heart is reversed (figure 1A, red arrow), as are the looping of the gut and position of the pancreas. The most conspicuous asymmetries in the CNS are found in the epithalamic region of the dorsal forebrain; in this region there is a left-sided nucleus of neurons called the parapineal and the left and right habenular nuclei show various asymmetries including their connections to other neurons (Figure 1B). Importantly, in fsi embryos with reversed heart looping the brain asymmetries are reversed concordantly (Figure 1).
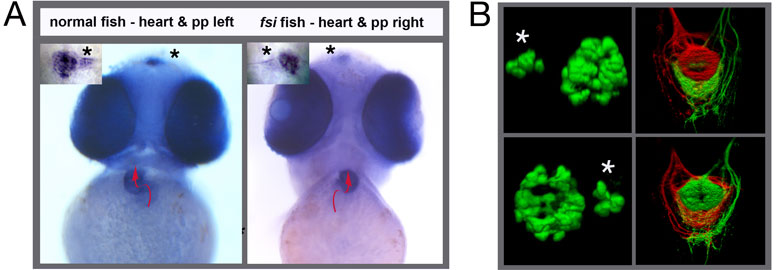
Figure 1. (A) In normal fish the heart loops from left to right and a small photoreceptive structure in the brain called the parapineal is located to the left side of the midline. Both asymmetries are reversed in the fsi fish line. The pictures show frontal views of young zebrafish embryos. A red arrow indicates the direction of heart looping and the asterisks point to the position of the small parapineal next to the larger pineal nucleus. (B) The left panels show 3-D reconstructions of the fluorescently labeled pineal and parapineal (asterisk) nuclei in normal (top) and reversed fsi fish (bottom). The top right panel depicts the projections of habenular neurons from the left side (red) and right side (green) to a target in the midbrain; in normal fish the left side sends projections to the top (dorsal) part of this target, the right side to the bottom (ventral) part. Again, the laterality of this asymmetry is reversed in fsi fry (bottom right panel)
Some behaviours are lateralised in zebrafish such as eye preference when viewing a conspecific or their own reflection. We wanted to know if this bias would be changed in larval fish (fry) that show a reversal of brain asymmetries. We tested fsi fry that showed a reversal of heart position (which we can see in living fry) and parapineal position (which is visible through expression of a fluorescent transgene in living fry; see Figure 1B left panels). For this test, we place fry into a tank with mirrored walls (Fig 2A; top) and score which eye the fry use to look at their reflection (Fig 2A; bottom). Interestingly, we found that indeed the characteristic eye switching and laterality of eye preference is reversed in fry with reversed CNS asymmetries.

Figure 2. fsi fry with reversed visceral and brain asymmetries show reversal of eye preference when viewing their own reflection. (A) The top panel shows a schematic of the mirror tank used in the test, the bottom panel how we score which eye is used by the fry for viewing. (B) Normal fry (LH, blue line) show a slight bias to use their right eye initially, but then switch to left eye use and back to left eye use. This bias is reversed in fsi fry with reversed asymmetries (RH, pink line).
Using a different test, the swimway, we examined if there is a bias in turning when fry enter a new compartment driven by their preference to move towards the light. Fry are place into a lit start compartment, the light is then switched to the next compartment and the time it takes for the fry to swim into the next compartment (latency) and the laterality of turning is measured. A slight leftward bias was found for the laterality of turning for both groups, normal and reversed (not shown). However, we did see a difference between the groups when we challenged fry by placing a novel object (black stripe) into successive compartments (Fig 3A). Reversed fsi fry appeared to be bolder when exploring spaces featuring novel visual cues; they emerged into successive compartments with less delay (latency) than their normal siblings (Fig 3B).
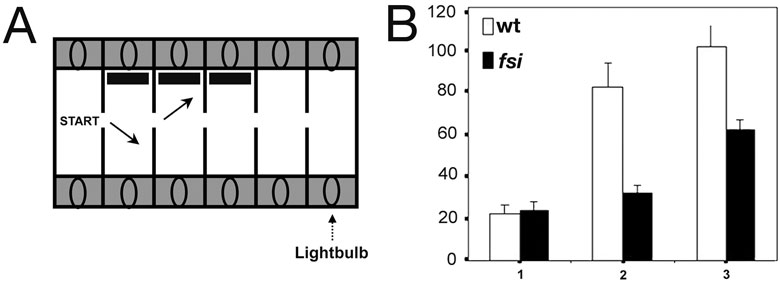
Figure 3. fsi fry with reversed asymmetries emerge into successive compartments with less delay than their normal siblings when encountering a novel visual object. (A) Fry are placed into the lit start compartment, and are then motivated to enter the next compartment by turning off the light in the start compartment and turning it on in the adjacent compartment. This is repeated for 3 successive compartments, each successive compartments feature a novel object (black stripe) on the side wall. (B) Fry are encountering a novel visual object upon emerging into the new compartment. Numbers 1, 2 and 3 on the horizontal axis show the number of consecutive compartments entered; the vertical axis shows the average delay of entering the next compartment after switching the light (in seconds).
Our findings support the idea that asymmetries in the brain are directly correlated to at least some lateralised behaviours. In addition, we found that fsi fry with reversed CNS asymmetries show a distinct behaviour compared to their normal siblings; they are less afraid to move into novel spaces, even if they encounter novel visual stimuli. This suggests that some lateralised behaviour is reversed in fry with reversed brain asymmetries and that brain reversal can lead to novel behaviours such as increased boldness. We are currently expanding these studies to include a wider range of behavioural tests.
Further information
McManus C. (2005) Reversed bodies, reversed brains, and (some) reversed behaviors: of zebrafish and men. Dev Cell. 8:796-7
Lin SY, Burdine RD (2005) Brain asymmetry: switching from left to right. Curr Biol. 15:R343-5
If you have questions about this work please visit our asymmetry research pages or contact Anukampa Barth or Steve Wilson
This study was a collaboration with Richard Andrew's group at the University of Sussex. Work from our lab on this study was supported by the Wellcome Trust.