
Matthias Carl and Steve Wilson
Original paper reference:
Wnt/Axin1/β-catenin signaling regulates asymmetric Nodal activation, elaboration, and concordance of CNS asymmetries
The vertebrate brain is an immensely complex structure which exhibits numerous neuroanatomical and functional asymmetries. In humans, some of these asymmetries are evident as biases in hand preference and from the different deficits that arise as a consequence of strokes on left or right sides of the brain. However, it is very difficult to study brain development in humans and so we use animal model systems to study the genetic and cellular mechanisms that underlie brain asymmetry.
Due in part to its small size, relative simplicity and transparency, the zebrafish embryo is particulary well suited to study the genetic pathways that allow the correct establishment of brain asymmetries. In the fish brain, several structures develop prominent asymmetries that are easy to visualise. In the dorsal part of the brain, so called parapineal neurons (nerve cells) are found exclusively on the left side of the brain and also the bilaterally positioned clusters of habenular neurons exhibit asymmetric features (figure 1).
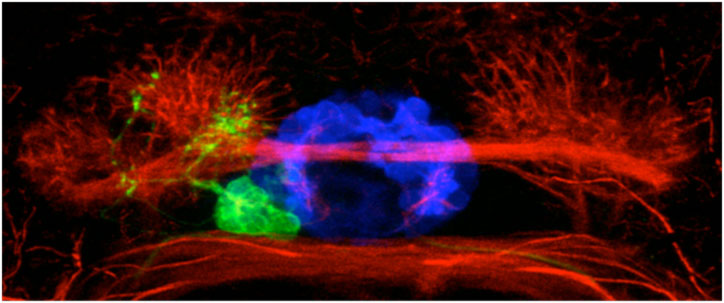
Figure 1. An image of asymmetric neurons in the larval fish brain. The blue neurons are at the midline of the brain, the green parapineal neurons are on the left and the red habenular neurons are differently organised on the left and the right.
In the search for genes that underlie the laterality/handedness of these brain asymmetries, we showed the Nodal signalling pathway plays an important and evolutionary conserved role. Nodal is one of a small number of genetic pathways that allow cells to communicate with each other during embryonic development. Some genes in this signalling cascade are active during a short period of time only on the left side of the brain and determine the subsequent directionality of the migration of parapineal cells from the midline towards the left side of the brain. Subsequent to this, further signals from the parapineal neurons are important to elaborate asymmetries in the habenulae. These various interactions eventually lead to the development of asymmetric neuronal circuits and behaviour.
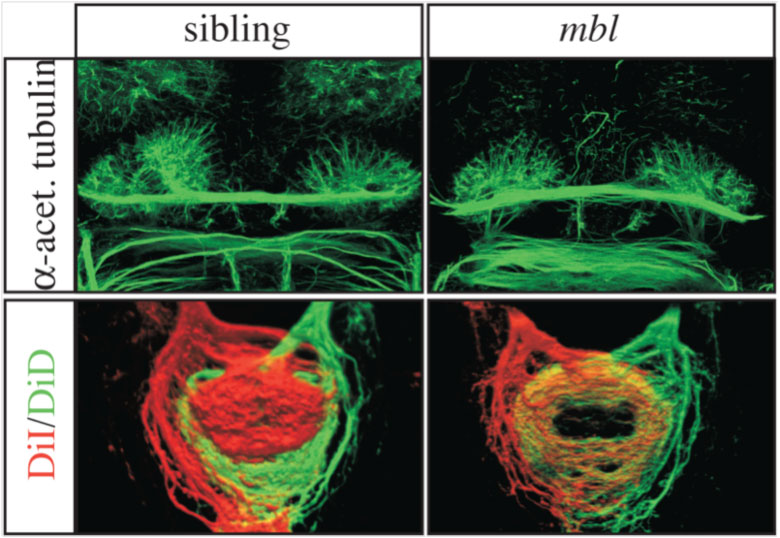
Figure 2. Images of brains of normal (sibling on the left) and masterblind/Axin1 (mbl on the right) mutant embryos. The habenulae in the top panel are asymmetric in the normal brain but symmetric in the mutant. (Bottom panel) These panels shows the connections made by the left habenula neurons (red) and the right habenula neurons (green). Left and right connections are segregated in the normal embryo but intermixed in the mutant consistent with the brain having symmetric double right character.
Although Nodal signalling in the left brain determines the direction of brain asymmetry, this begs the question of how Nodal signalling is restricted to the left side of the brain in the first place? In our search for genes which influence asymmetric Nodal signalling, we found a zebrafish mutant in which the asymmetry of Nodal is lost and the brains develop symmetrically, as if they are double right-sided (figure 2). These masterblind mutants carry a mutation in Axin1, a gene which acts to turn down the activity of another signalling cascade, the Wnt/β-catenin pathway. We were able to show that Axin1 mediated signaling during very early stages of embryonic development in the future brain is required to establish Nodal gene asymmetry. Too much Wnt/β-catenin signalling at these stages (as happens in the mutant) leads to activation of Nodal signalling on both sides of the brain.
These and other results led us to propose a rather complicated and counter-intuitive model of how Nodal comes to be on the left. A first step is that both sides of the brain are primed to express Nodal but the pathway is switched off on both sides (this step involves the Wnt pathway). Later in development, a signal from the left side of the body signals to the brain to remove the 'off-switch' and consequently Nodal is activated only in the left brain.
So we take a step backwards and now ask why does only the left side of the body send the signal to the brain. It turns out that the Nodal pathway is involved again. It is required to determine the asymmetries in our internal organs like the heart, liver and lungs and in doing this, the pathway appears to have been co-opted to signal left from right in the brain.
Our studies also revealed a later role for Wnt signalling in the communication between the asymmetric brain structures and this is a topic that we are currently following up.
Here's a link to our research description pages.
A related study from Lila Solnica-Krezel's group was published together with ours
Inbal A, Kim SH, Shin J, Solnica-Krezel L. (2007)
Six3 represses nodal activity to establish early brain asymmetry in zebrafish
Neuron 55:407-15
And both studies are reviewed in
Sagasti A (2007)
Three ways to make two sides: genetic models of asymmetric nervous system development
Neuron. 55:345-51
If you have any more questions about this work, please contact Matthias or Steve Wilson
Our work on this project was primarily supported by the BBSRC and Wellcome Trust with some additional funding from the EU.