

The retina Is composed of the neural retina and retinal pigmented epithelium. This multilayered structure conveys visual information to the brain.
You can explore the different regions of the zebrafish retina and overlying ectodermal structures, the lens and cornea by selecting one of the images below. The specific layers and structures referenced in this tutorial are highlighted in white. You can click on an image to see which transgenic lines or antibodies label each specific layer or the neural retina. For a more general overview of the neuroanatomy of the zebrafish retina please read on.
Retinal Regions









The Eye
The eye is a compound organ that functions as the sensory apparatus of the visual system. It detects information in the form of refracted light and together with visual centres in the brain it provides visual perception of the surrounding environment.The eye is composed of the retina, optic nerve, lens, cornea and iris. These structures derive from three types of tissue during embryogenesis: neural ectoderm, extraocular mesenchyme and surface ectoderm. The neural ectoderm gives rise to the retinal derivatives (neural retina, ciliary margin zone (CMZ), retinal pigmented epithelium (RPE), and optic stalk/nerve); the extraocular mesoderm gives rise to the corneal endothelium, iris, and other extraocular structures; the surface ectoderm gives rise to the lens and the corneal epithelium. For the sake of this neuroanatomical atlas, we concentrate on the parts of the eye that are derived from the neural plate, which evaginate from the region of the forebrain called the diencephalon.
The neural Retina
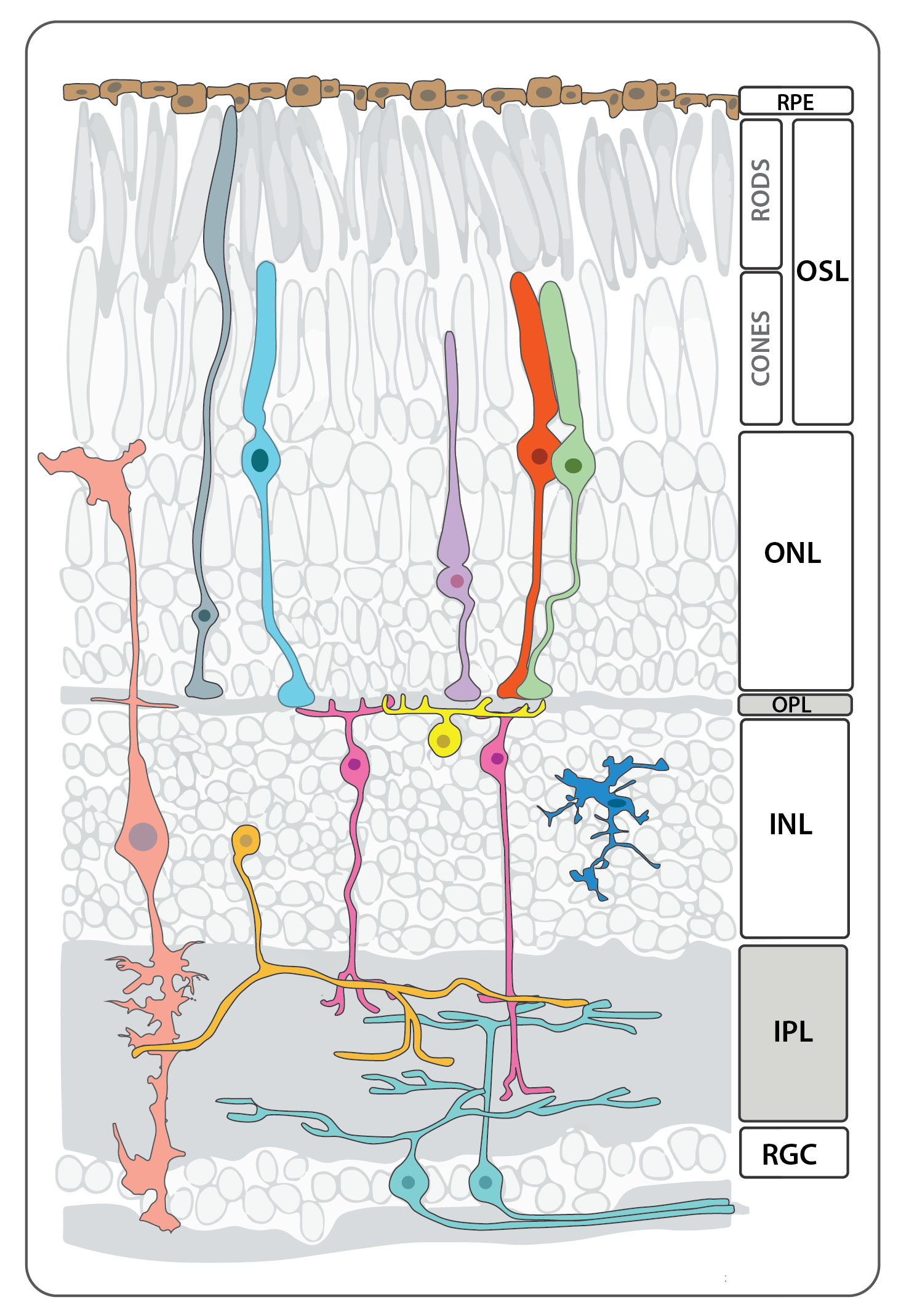
Schematic depicting the different cell types and layers that make up the neural retina in an adult zebrafish adapted from Baden et al., 2019.
Vision, one of the most important senses, begins when a photon strikes opsin molecules located in the photoreceptor outersegments. Photoreceptors translate this physical stimulus into an electrochemical signal that is transmitted and refined through the interneurons of the retina. Within milliseconds, this message converges on the retinal ganglion neurons that retinotopically project their axons to specific locations in the optic tectum, the visual processing center of the zebrafish brain. These retinotopic projections preserve the nasal-temproal and dorsal-ventral spatial resolution of the image detected by the eye.
Like most classes of extant vertebrates, the zebrafish neural retina is composed of six major cell types, five types of neurons and a single glial cell, the Müller glia. The cells are arranged in three basic layers: the innermost ganglion cell layer, inner nucler layer, and the outer photoreceptor layer, and these are separated by two intervening layers of synapses, the inner and outer plexiform layers. The cell bodies of the rod and cone photoreceptors reside in the outer nuclear layer while those of bipolar cells, horizontal cells, and amacrine cells, as well as the Muller glia, are found in the inner nuclear layer. All retinal ganglion cell bodies, along with those of displaced amacrine cells occupy the ganglion cell layer. The five major classes of neurons can be subdivided into numerous subpopulations based on morphological, immunohistochemical and physiological profiles, and a more detailed description of the immunolabeling patterns for many of these subpopulations and structures can be found in Yazulla and Studholme, 2001.
The neural retina is composed of the following general types of cells, starting from apical to basal:
Retinal outer-nuclear layer cell types:
Photoreceptors
Photoreceptor cells span the outer nuclear layer as well as the outer and inner segment layers. The light sensing cells of the retina, the cone and rod photoreceptors, are located in the apical-most layer of the retina, and display a stereotypical subcellular organization with their nuclei basal to the inner segment (cell body or soma) and outer segment, which abuts the retinal pigmented epithelium (RPE) and is full of membrane invaginations packed with light-sensitive cell-specific opsins. All photoreceptors have opsin-containing outersegments, which are replaced on a daily basis. The older membrane and protein debris is shed from the distal tip of the outersegment and phagocytosed by the adjacent RPE cells. Because zebrafish are diurnal, their retinae contains a large number of bright-light sensitive cone subtypes in addition to dim-light sensitive rod photoreceptors, all of which are organized into regular mosaic patterns that can best be visualized in tangential sections. Below is more detailed information about the distinct types of photoreceptors found in zebrafish.


Retinal Inner-nuclear layer cell types:



Retinal ganglion cell Layer

Retinal Glia


Retinal Pigmented Epithelium (Non-Neural Retina)






CLick button below to read more on Eye Development in zebrafish
Author: Kara Cerveny
Key Publications
Robles, E., Laurell, E., Baier, H. (2014)
The Retinal Projectome Reveals Brain-Area-Specific Visual Representations Generated by Ganglion Cell Diversity.
Current biology : CB. 24(18):2085-96.
Almeida, A.D., Boije, H., Chow, R.W., He, J., Tham, J., Suzuki, S.C., Harris, W.A. (2014)
Spectrum of Fates: a new approach to the study of the developing zebrafish retina.
Development (Cambridge, England). 141:1971-80.