Forebrain > olfactory epithelium

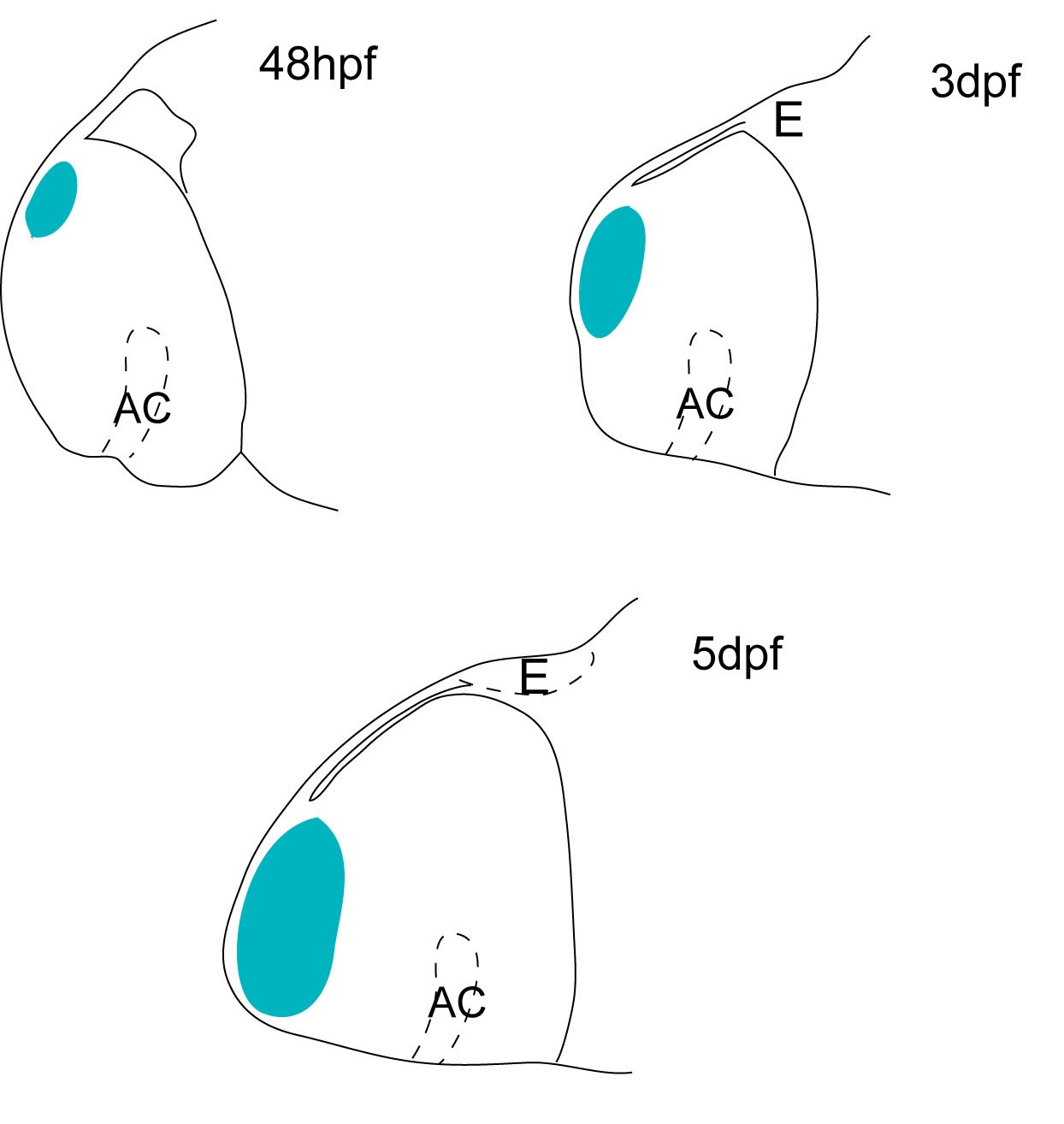
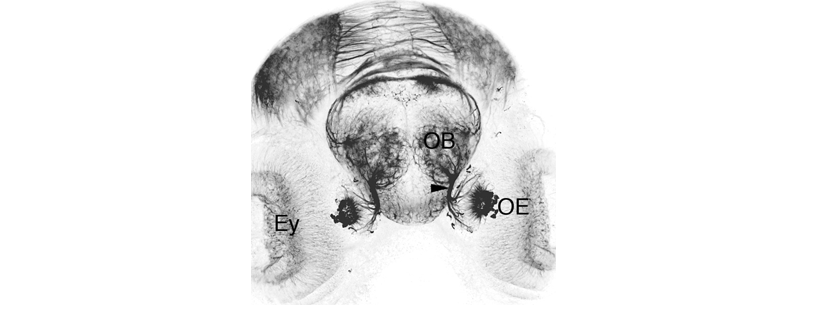
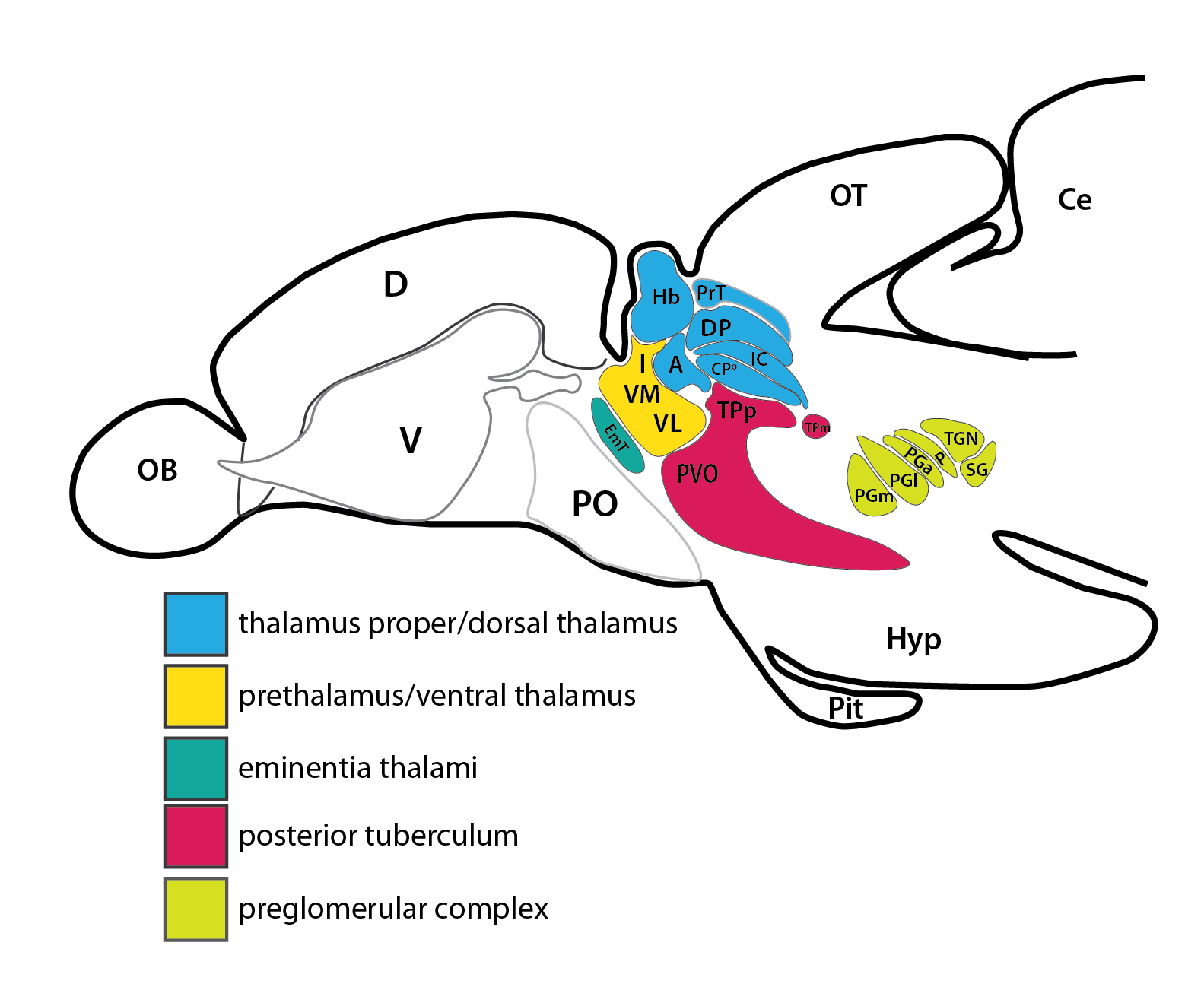
Schematic showing the position of the olfactory epithelium in sagittal, horizontal and coronal sections through the zebrafish brain. Based on the anatomical segmentation of 3 day old zebrafish larval brain by Thomas Müller, Olaf Ronneberger, Wolfgang Driever and colleagues. For details see Ronneberger et al., Nat. Meth. 2012 and http://vibez.informatik.uni-freiburg.de
Abbreviations: Ce, cerebellar plate; D, dorsal telencephalon/pallium; E, epiphysis; EmT, eminentia thalami; Hb, habenula; Hyp, hypothalamus; MO, medulla oblongata; OB, olfactory bulb; OT, optic tectum; PG, preglomerular complex; PO, preoptic area;PrT, pretectum; PTd, posterior tuberculum dorsal part; PTh, prethalamus; PTv posterior tuberculum ventral part; Teg, tegmentum; Th, thalamus; TS, torus semicircularis; V, ventral telencephalon/subpallium; Va, valvula cerebelli.
Description
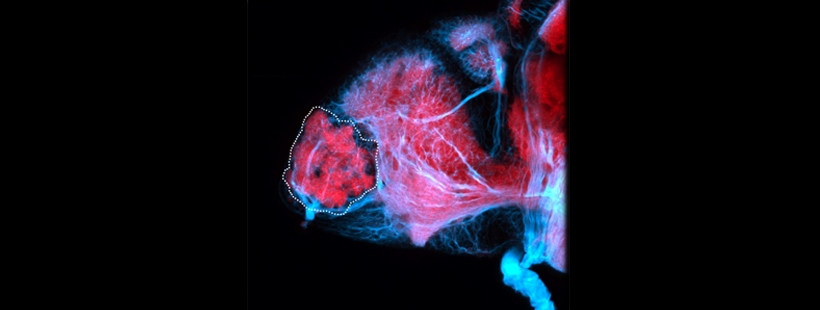
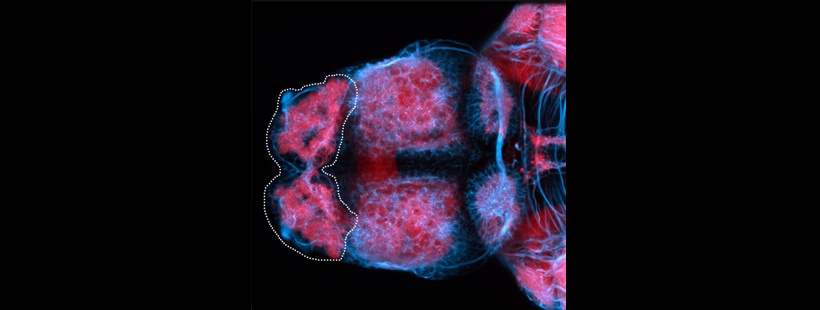
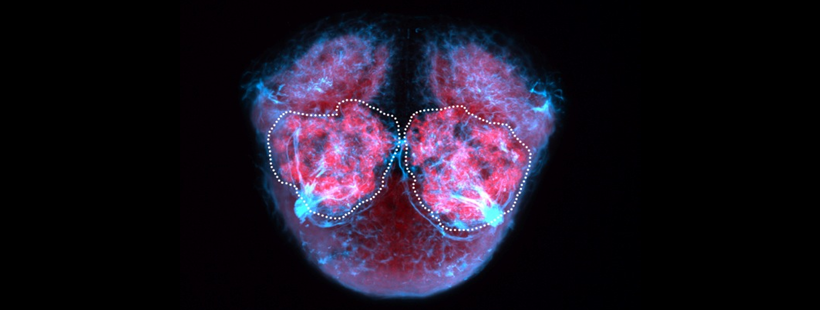
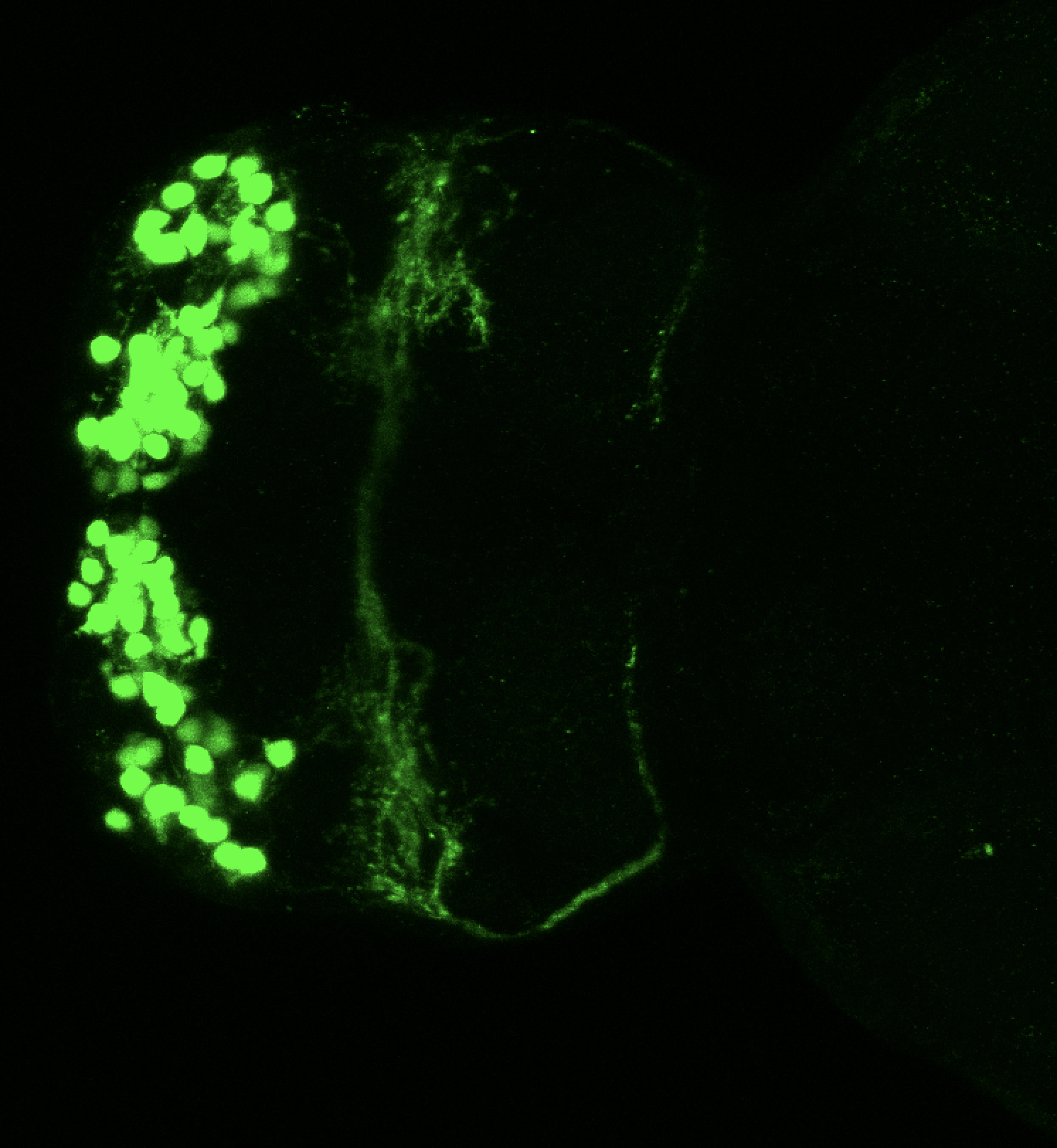
Also known as the nasal placode. The olfactory epithelium contains the cell bodies of olfactory sensory neurons that project axons into the olfactory bulb where they contact mitral cells at olfactory bulb glomeruli. In zebrafish the predominant subtypes are ciliated sensory neurons that have long dendrites and microvillous sensory neurons, which have short dendrites, these cells are placodally derived (Aguillon et al., 2018). A third group of neurons associated with the olfactory epithelium express gonadotropin releasing hormone 3 (Gnrh3), these may be the cell bodies of the terminal nerve cranial nerve 0.
Ontology
is part of: olfactory system
has parts:
OLS Tree Diagram

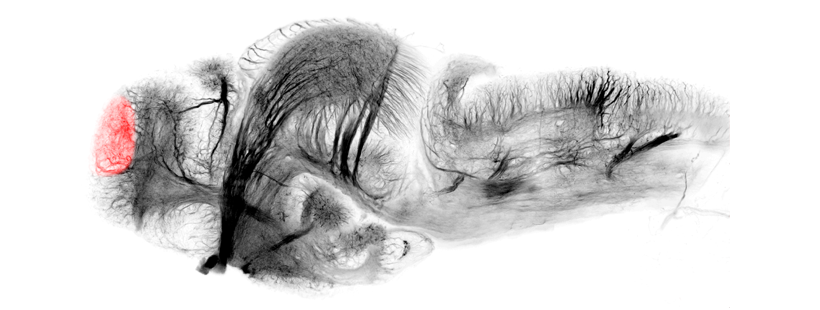
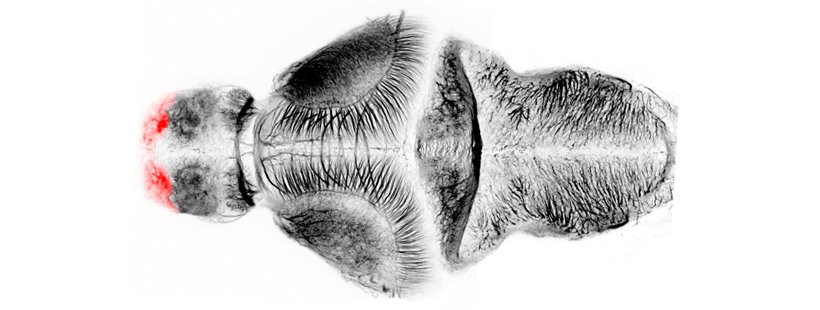
Anatomical segmentation of 3 day old zebrafish larval brain by Thomas Müller, Olaf Ronneberger, Wolfgang Driever and colleagues. For details see Ronneberger et al., Nat. Meth. 2012 and http://vibez.informatik.uni-freiburg.de
Transgenic Lines that label this brain region



Antibodies that label this brain region



Key Publications
Anne Hansen, Zeiske Eckart
The Peripheral Olfactory Organ of the Zebrafish, Danio rerio: an Ultrastructural Study
Chemical Senses, Volume 23, Issue 1, February 1998, Pages 39-48, https://doi.org/10.1093/chemse/23.1.39
Kathleen E. Whitlock and Monte Westerfield
A Transient Population of Neurons Pioneers the Olfactory Pathway in the Zebrafish
Journal of Neuroscience 1 November 1998, 18 (21) 8919-8927; DOI: https://doi.org/10.1523/JNEUROSCI.18-21-08919.1998
Raphaël Aguillon, Julie Batut, Arul Subramanian, Romain Madelaine, Pascale Dufourcq, Thomas F Schilling, Patrick Blader
Cell-type heterogeneity in the early zebrafish olfactory epithelium is generated from progenitors within preplacodal ectoderm
eLife 2018;7:e32041 DOI: 10.7554/eLife.32041
Back to Forebrain